Katherine Hayles distinguishes three phases in the history of cybernetics and highlights how each successive development continues a “tendency to reconfigure boundaries” (Mitchell & Hansen, 2010, pp. 147, pp. 149):
Third-order cybernetics redraws the boundary once again to locate both the observer and the system within complex, networked, adaptive, and coevolving environments through which information and data are pervasively flowing (Mitchell & Hansen, 2010, p. 149).
I am interested in these shifting boundaries, particularly when they stimulate dialectical views on human identity – where do we begin, where do we end – with the implication that these never have simple definitive answers. Substitute when to where, and the questions are quickly overloaded with political tensions, raising dual issues about beginning and end of life. But they are just as powerful when viewed causally and functionally.
For instance, Gary Schneider explores his identity as human and multi-cellular animal, in a series of Genetic Self Portraits. They span micro to macro scales, including chromosomes, a cell from the lining of the cheeks, a hair, or an iris:
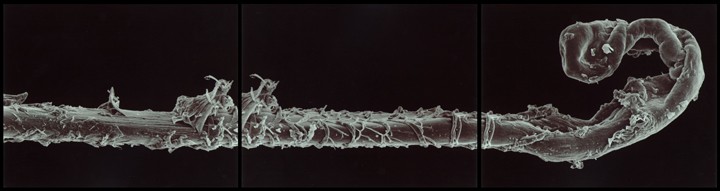
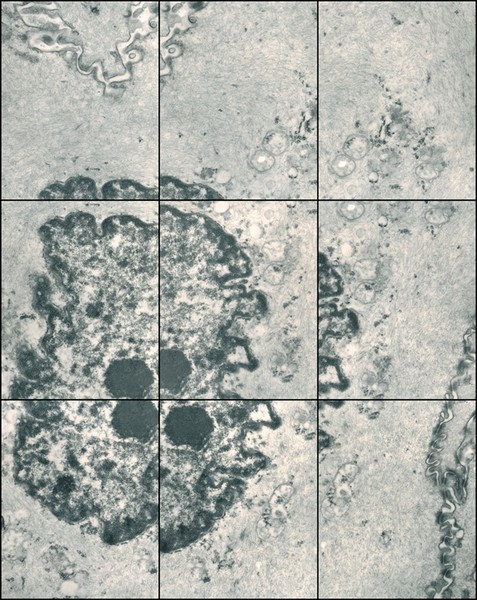
In 1999, Schneider complemented this series with new photos of his intestinal flora, cells from an altogether different lineage. Bacteria are prokaryotes: they have no nucleus, mitochondria or other organelles; unlike eukaryote cells from which all plants, animals and fungi are composed. While prokaryotes might seem primitive, they are really the chemists of the natural world, with a vast repertoire of potential chemical reactions unlike anything animal or plant cells can do.
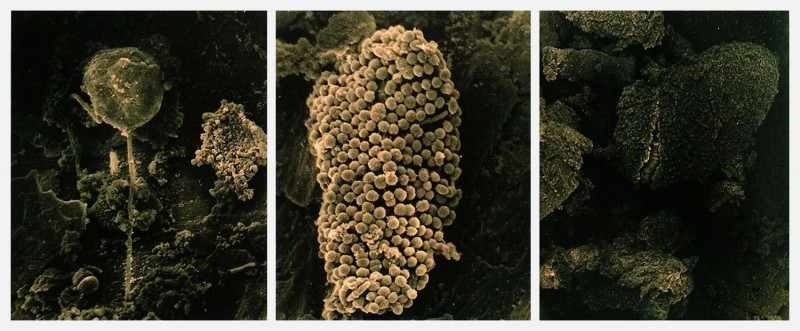
Edgar Lissel‘s work, which he calls Microbial Art, develops a similar gesture and provides a great bridge from biomedia to cybernetics. He combines photography and microorganisms to create living images by exposing petri dishes filled with nutrients and light-seeking cyanobacteria, a biological film instead of photographic paper. Note that microbiologists do talk about biofilms (the slimy mats created by bacterial colonies), a fortuitous lexical convergence.

The microorganisms here are occasionally the subjects (in the second image for instance). But more importantly they provide the materiality out of which the images come together. And note that this materiality is not simply a case of static pigments. The images manifest behaviors: the cyanobacteria moving and seeking light.
A more recent series of pieces figures decidedly human shapes: imprints of an arm, a hand, feet:
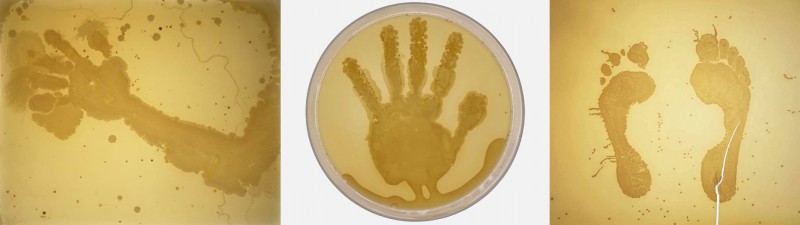
These were created by dipping body parts into a sterile nutrient solution. The bacterial growths are from the artist’s own microbiota, seeded simply through contact. The pieces biologically render the shape of the human body through its symbiotes and commensals: the non-us which permeates us and makes us possible.
Maturana and Varela’s concept of autopoïesis was initially conceived at the cellular level. It is a powerful concept: defining a system by its ability to sustain and duplicate itself. But as we trace our operational closure – that boundary within which our processes reflexively define themselves – we are led to expand it in multiple ways. Reaching outwardly reveals “the observer [as] constructed within social and linguistic environments” (Mitchell & Hansen, 2010, pp. 149).
But complexity also lies inwardly at the micro-scale. The images so far have gone in magnification up to the cell as a unit. But within this unit lies yet another complex system and environment. The following animation was developed by xvivo for Harvard University, and shows one eukaryote cell as a vast landscape bustling with activity.
It seems wherever we look, at every scale, we find “complex, networked, adaptive, and coevolving environments through which information and data are
pervasively flowing” (Mitchell & Hansen, 2010, pp. 149).
Leave me a Comment
You must be logged in to post a comment.